





北京大學地質數字博物館
地質的演化歷程
生命的起源
生命的起源應當追溯到與生命有關的元素及化學分子的起源.因而,
生命的起源過程應當從宇宙形成之初、通過所謂的“大爆炸”產生了碳、氫、氧、氮、磷、硫等構成生命的主要元素談起。
大約在66億年前,銀河系內發生過一次大爆炸,其碎片和散漫物質經過長時間的凝集,
大約在46億年前形成了太陽系。作為太陽系一員的地球也在46億年前形成了。
接著,冰冷的星雲物質釋放出大量的引力勢能,再轉化為動能、熱能,致使溫度升高,
加上地球內部元素的放射性熱能也發生增溫作用,故初期的地球呈熔融狀態。
高溫的地球在旋轉過程中其中的物質發生分異,重的元素下沉到中心凝聚為地核,
較輕的物質構成地幔和地殼,逐漸出現了圈層結構。
這個過程經過了漫長的時間,大約在38億年前出現原始地殼,這個時間與多數月球表面的岩石年齡一致。
生命的起源與演化是和宇宙的起源與演化密切相關的。
生命的構成元素如碳、氫、氧、氮、磷、硫等是來自“大爆炸”後元素的演化。
資料表明前生物階段的化學演化並不局限於地球,在宇宙空間中廣泛地存在著化學演化的產物。
在星際演化中,某些生物單分子,如氨基酸、嘌呤、嘧啶等可能形成於星際塵埃或凝聚的星雲中,
接著在行星表面的一定條件下產生了象多肽、多聚核苷酸等生物高分子。
通過若干前生物演化的過渡形式最終在地球上形成了最原始的生物系統,即具有原始細胞結構的生命。
至此,生物學的演化開始,直到今天地球上產生了無數複雜的生命形式。
38億年前,地球上形成了穩定的陸塊,各種證據表明液態的水圈是熱的,甚至是沸騰的。
現生的一些極端嗜熱的古細菌和甲烷菌可能最接近於地球上最古老的生命形式,其代謝方式可能是化學無機自養。
澳大利亞西部瓦拉伍那群中35億年前的微生物可能是地球上最早的生命證據。
原始地殼的出現,標誌著地球由天文行星時代進入地質發展時代,具有原始細胞結構的生命也開始逐漸形成。
但是在很長的時間內尚無較多的生物出現,一直到距今5.4億年前的寒武紀,帶殼的後生動物才大量出現,
故把寒武紀以後的地質時代稱為顯生宙
藻類和無脊椎動物時代
前寒武紀
太古宙(Archean)是最古老的地史時期。
從生物界看,這是原始生命出現及生物演化的初級階段,當時只有數量不多的原核生物,他們只留下了極少的化石記錄。
從非生物界看,太古宙是一個地殼薄、地熱梯度陡、火山—岩漿活動強烈而頻繁、
岩層普遍遭受變形與變質、大氣圈與水圈都缺少自由氧、形成一系列特殊沉積物的時期;
也是一個矽鋁質地殼形成並不斷增長的時期,又是一個重要的成礦時期。
元古宙(Proterozoic)初期地表已出現了一些範圍較廣、厚度較大、相對穩定的大陸板塊。
因此,在岩石圈構造方面元古代比太古代顯示了較為穩定的特點。
早元古代晚期的大氣圈已含有自由氧,而且隨著植物的日益繁盛與光合作用的不斷加強,大氣圈的含氧量繼續增加。
元古代的中晚期藻類植物已十分繁盛,明顯區別於太古代。
震旦紀(Sinian period)是元古代最後期一個獨特的地史階段。
從生物的進化看,震旦系因含有無硬殼的後生動物化石,而與不含可靠動物化石的元古界有了重要的區別;
但與富含具有殼體的動物化石的寒武紀相比,震旦系所含的化石不僅種類單調、數量很少而且分佈十分有限。
因此,還不能利用其中的動物化石進行有效的生物地層工作。
震旦紀生物界最突出的特徵是後期出現了種類較多的無硬殼後生動物,末期又出現少量小型具有殼體的動物。
高級藻類進一步繁盛,微體古植物出現了一些新類型,疊層石在震旦紀早期趨於繁盛,後期數量和種類都突然下降。
再從岩石圈的構造狀況來看,震旦紀時地表上已經出現幾個大型的、相對穩定的大陸板塊,
之上已經是典型的蓋層沉積,與古生界相似。因此,震旦紀可以被認為是元古代與古生代之間的一個過渡階段。
寒武紀 三葉蟲時代
寒武紀(Cambrian period)是古生代的第一個紀,開始於距今5.4億年,延續了4000萬年。
寒武紀是生物界第一次大發展的時期,當時出現了豐富多樣且比較高級的海生無脊椎動物,
保存了大量的化石,從而有可能研究當時生物界的狀況,並能夠利用生物地層學方法來劃分和對比地層,
進而研究有機界和無機界比較完整的發展歷史。
比較著名的有早寒武世雲南的澄江動物群、加拿大中寒武世的布林吉斯葉岩生物群。
寒武紀的生物界以海生無脊椎動物和海生藻類為主。
無脊椎動物的許多高級門類如節肢動物、棘皮動物、軟體動物、腕足動物、筆石動物等都有了代表。
其中以節肢動物門中的三葉蟲綱最為重要,其次為腕足動物。
此外,古杯類、古介形類、軟舌螺類、牙形刺、鸚鵡螺類等也相當重要。
拋開牙形石不說,高等的脊索動物還有許多其他代表,
如我國雲南澄江動物群中的華夏鰻、雲南魚、海口魚等,加拿大布林吉斯葉岩中的皮開蟲,美國上寒武統的鴨鱗魚。
|
|
|
|
|
|
|
| 腔腸動物 | 布林吉斯動物群(Ayshea) | 水母狀動物 | 節肢動物(Marrella) | 節肢動物(Leanchoilia) | 節肢動物 |
奧陶紀 原始的脊椎動物出現
奧陶紀(Ordovician period)是古生代的第二個紀,開始於距今5億年,延續了6500萬年。
奧陶紀是地史上海侵最廣泛的時期之一。
在板塊內部的地台區,海水廣布,表現為濱海淺海相碳酸鹽岩的普遍發育,在板塊邊緣的活動地槽區,
為較深水環境,形成厚度很大的淺海、深海碎屑沉積和火山噴發沉積。
奧陶紀末期曾發生過一次規模較大的冰期,其分佈範圍包括非洲,特別是北非、南美的阿根廷、
玻利維亞以及歐洲的西班牙和法國南部等地。
奧陶紀的生物界較寒武紀更為繁盛,海生無脊椎動物空前發展,其中以筆石、三葉蟲、鸚鵡螺類和腕足類最為重要,
腔腸動物中的珊瑚、層孔蟲,棘皮動物中的海林檎、海百合,節肢動物中的介形蟲,苔蘚動物等也開始大量出現。
奧陶紀中期,在北美洛磯山脈地區出現了原始脊椎動物異甲魚類——星甲魚和顯褶魚,
在南半球的澳大利亞也出現了異甲魚類。植物仍以海生藻類為主。
 |
 |
 |
 |
 |
 |
| 蠕形石蝦(Helminthochiton) | 直角石(Orthoceras) | 弓形角石(Cyrtoceras) | 休倫角石(Huronia) | 肋棘海林檎(Pleurocystes) | 肋棘海林檎(Pleurocystes) |
粿蕨植物和魚類時代
志留紀 筆石的時代,陸生植物和有頜類出現
志留紀(Silurian period)是早古生代的最後一個紀。
本紀始於距今4.35億年,延續了2500萬年。由於志留系在波羅的海哥德蘭島上發育較好,因此曾一度被稱為哥德蘭系。
志留系三分性質比較顯著。
一般說來,早志留世到處形成海侵,中志留世海侵達到頂峰,晚志留世各地有不同程度的海退和陸地上升,
表現了一個巨大的海侵旋回。志留紀晚期,地殼運動強烈,古大西洋閉合,一些板塊間發生碰撞,
導致一些地槽褶皺升起,古地理面貌巨變,大陸面積顯著擴大,生物界也發生了巨大的演變,
這一切都標誌著地殼歷史發展到了轉折時期。
志留紀的生物面貌與奧陶紀相比,有了進一步的發展和變化。
海生無脊椎動物在志留紀時仍占重要地位,但各門類的種屬更替和內部組分都有所變化。
如筆石動物保留了雙筆石類,新興的單筆石類也很繁盛;
腕足動物內部的構造變得比較複雜,如五房貝目、石燕貝目、小嘴貝目得到了發展;
軟體動物中頭足綱、鸚鵡螺類顯著減少,而雙殼綱、腹足綱則逐步發展;
三葉蟲開始衰退,但蛛形目和介形目大量發展;
節肢動物中的板足鱟,也稱“海蠍”在晚志留世海洋中廣泛分佈;
珊瑚綱進一步繁盛;棘皮動物中海林檎類大減,海百合類在志留紀大量出現。
脊椎動物中,無頜類進一步發展,有頜的盾皮魚類和棘魚類出現,這在脊椎動物的演化上是一重大事件,
魚類開始征服水域,為泥盆紀魚類大發展創造了條件。
植物方面除了海生藻類仍然繁盛以外,晚志留世末期,陸生植物中的裸蕨植物首次出現,
植物終於從水中開始向陸地發展,這是生物演化的又一重大事件。
 |
 |
 |
 |
 |
 |
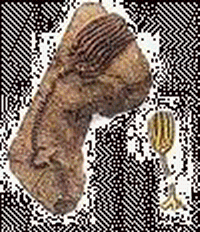
| 寬角螺(Platyceras) | 細網筆石(reliortes) | 全臍螺(Euomphalus) | 雙股海百合(Dimerocrinites) | 神螺(Bellerophon) | 星苔蘚蟲(Constellania) |
泥盆紀 魚類的時代
泥盆紀(Devonian period)是晚古生代的第一個紀,開始於距今4.1億年,延續了約5500萬年。
泥盆紀古地理面貌較早古生代有了巨大的改變。表現為陸地面積的擴大,陸相地層的發育,生物界的面貌也發生了巨大的變革。
陸生植物、魚形動物空前發展,兩棲動物開始出現,無脊椎動物的成分也顯著改變。
腕足類在泥盆紀發展迅速,志留紀開始出現的石燕貝目成為泥盆紀的重要化石。
此外,穿孔貝目、扭月貝目、無洞貝目和小嘴貝目在劃分和對比泥盆紀地層中也極為重要。
泡沫型和雙帶型四射珊瑚相當繁盛。早泥盆世以泡沫型為主,雙帶型珊瑚開始興起;
中、晚泥盆世以雙帶型珊瑚占主要地位。
鸚鵡螺類大大減少,菊石中的棱菊石類和海神石類繁盛起來。
正筆石類大部分絕滅,早泥盆世殘存少量單筆石科的代表。
竹節石類始於奧陶紀,泥盆紀一度達到最盛,泥盆紀末期絕滅。
其中以薄殼型的塔節石類最繁盛,光殼節石類也十分重要。
牙形石演化到泥盆紀又進入一個發展高峰,這個時期以平臺型分子大量出現為特徵。
昆蟲類化石最早也發現於泥盆紀。
泥盆紀是脊椎動物飛越發展的時期,魚類相當繁盛,各種類別的魚都有出現,故泥盆紀被稱為 “魚類的時代”。
早泥盆世以無頜類為多,中、晚泥盆世盾皮魚相當繁盛,它們已具有原始的顎,偶鰭發育,成歪形尾。
早泥盆世裸蕨植物較為繁盛,有少量的石鬆類植物,多為形態簡單、個體不大的草本類型;
中泥盆世裸蕨植物仍佔優勢,但原始的石鬆植物更發達,出現了原始的楔葉植物和最原始的真蕨植物;
晚泥盆世到來時,裸蕨植物瀕於滅亡,石鬆類繼續繁盛,節蕨類、原始楔葉植物獲得發展,新的真蕨類和種子蕨類開始出現。
 |
 |
 |
 |
 |
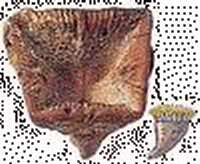 |
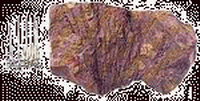
| 水角石(Hydnoceras) | 大葉藻(Zosterophyllum) | 亨頓蟲(Huntonia) | 杯海百合(Cyathocrinites) | 海神石(Clymenia) | 方錐珊瑚(Goniophyllum) |
蕨類植物和兩棲類動物時代
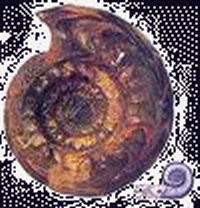
石炭紀 兩棲動物的時代
石炭紀(Carboniferous period)開始於距今約3.55億年至2.95億年,延續了6000萬年。
石炭紀時陸地面積不斷增加,陸生生物空前發展。
當時氣候溫暖、濕潤、沼澤遍佈,大陸上出現了大規模的森林,給煤的形成創造了有利條件。
石炭紀又是地殼運動非常活躍的時期,因而古地理的面貌有著極大的變化。
這個時期氣候分異現象又十分明顯,北方古大陸為溫暖潮濕的聚煤區,
岡瓦納大陸卻為寒冷的大陸冰川沉積環境。氣候分帶導致了動、植物地理分區的形成。
石炭紀的海生無脊椎動物與泥盆紀比較起來,有了顯著的變化。淺海底棲動物中仍以珊瑚、腕足類為主。
早石炭世晚期的浮游和游泳的動物中,出現了新興的筳類,菊石類仍然繁盛,三葉蟲到石炭紀已經大部分絕滅,只剩下幾個屬種。
最早發現於泥盆紀的昆蟲類,在石炭紀得到進一步的繁盛,已知石炭、二疊紀的昆蟲就達1300種以上。
陸生脊椎動物進一步繁盛,兩棲動物占到了統治地位。
早石炭世一開始,兩棲動物蓬勃發展,主要出現了堅頭類(也稱迷齒類),同時繁盛的還有殼椎類。
早石炭世的植物面貌與晚泥盆世相似,古蕨類植物延續生長,但只能適應於濱海低地的環境;
晚石炭世植物進一步發展,除了節蕨類和石鬆類外,真蕨類和種子蕨類也開始迅速發展。
裸子植物中的苛達樹是一種高大的喬木,成為造煤的重要材料之一。
 |
 |
 |
 |
 |
 |
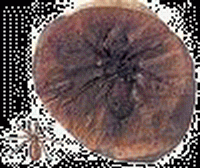
| 爪海膽(Melonechinus) | 星百合(Aclinocrinites) | 格雷伊夫蟲(Graeophonus) | 腹菊石(Gastrioceras) | 雷用諾角石(Rayonnoceras) | 原生木賊(Equisetites) |
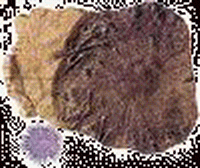
二疊紀 重要的成煤期
二疊紀(Permian period)是古生代的最後一個紀,也是重要的成煤期。
二疊紀開始於距今約2.95億年,延至2.5億年,共經歷了4500萬年。
二疊紀的地殼運動比較活躍,古板塊間的相對運動加劇,世界範圍內的許多地槽封閉並陸續地形成褶皺山系,
古板塊間逐漸拚接形成聯合古大陸(泛大陸)。陸地面積的進一步擴大,海洋範圍的縮小,自然地理環境的變化,
促進了生物界的重要演化,預示著生物發展史上一個新時期的到來。
二疊紀是生物界的重要演化時期。
海生無脊椎動物中主要門類仍是筳類、珊瑚、腕足類和菊石,但組成成分發生了重要變化。
節肢動物的三葉蟲只剩下少數代表,腹足類和雙殼類有了新的發展。
二疊紀末,四射珊瑚、橫板珊瑚、筳類、三葉蟲全都絕滅;腕足類大大減少,僅存少數類別。
脊椎動物在二疊紀發展到了一個新階段。
魚類中的軟骨魚類和硬骨魚類等有了新發展,軟骨魚類中出現了許多新類型,軟骨硬鱗魚類迅速發展。
兩棲類進一步繁盛。爬行動物中的杯龍類在二疊紀有了新發展;中龍類游泳於河流或湖泊中,以巴西和南非的中龍為代表;
盤龍類見於石炭紀晚期和二疊紀早期;獸孔類則是二疊紀中、晚期和三疊紀的似哺乳爬行動物,世界各地皆有發現。
早二疊世的植物界面貌與晚二疊世相似,仍以節蕨、石鬆、真蕨、種子蕨類為主。
晚二疊世出現了銀杏、蘇鐵、本內蘇鐵、松柏類等裸子植物,開始呈現中生帶的面貌。
 |
 |
 |
 |
 |
 |
| 原始海膽(Archaeocidaris) | 瓣齒魚(Petalodus) | 古鱈(Palaeoniscus) | 盜首螈(Diplocaulus) | 威爾金蛤(Wilkingia) | 基龍(Edaphosaurus) |
祼子植物和爬行動物時代
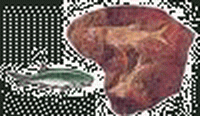
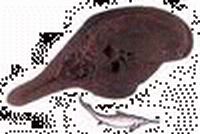
三疊紀 爬行動物和裸子植物的崛起
三疊紀(Triassic period)是中生代的第一個紀。始於距今2.5億年至2.03億年,延續了約5000萬年。
海西運動以後,許多地槽轉化為山系,陸地面積擴大,地台區產生了一些內陸盆地。
這種新的古地理條件導致沉積相及生物界的變化。從三疊紀起,陸相沉積在世界各地,
尤其在中國及亞洲其他地區都有大量分佈。古氣候方面,三疊紀初期繼承了二疊紀末期乾旱的特點;
到中、晚期之後,氣候向濕熱過渡,由此出現了紅色岩層含煤沉積、旱生性植物向濕熱性植物發展的現象。
植物地理區也同時發生了分異。
生物變革方面,陸生爬行動物比二疊紀有了明顯的發展。
古老類型的代表(如無孔亞綱和下孔亞綱)基本絕滅,新類型大量出現,並有一部分轉移到海中生活。
原始哺乳動物在三疊紀末期也出現了。
由於陸地面積的擴大,淡水無脊椎動物發展很快,海生無脊椎動物的面貌也為之一新。
菊石、雙殼類、有孔蟲成為劃分與對比地層的重要門類,而筳及四射珊瑚則完全絕滅。
爬行動物在三疊紀崛起,主要由槽齒類、恐龍類、似哺乳的爬行類組成。
典型的早期槽齒類表現出許多原始的特點,且僅限於三疊紀,其總體結構是後來主要的爬行動物以至於鳥類的祖先模式;
恐龍類最早出現於晚三疊世,有兩個主要類型:
較古老的蜥臀類和較進化的鳥臀類。
海生爬行類在三疊紀首次出現,由於適應水中生活,其體形呈流線式,四肢也變成槳形的鰭;
似哺乳爬行動物亦稱獸孔類,四肢向腹面移動,因此更適於陸地行走。
原始的哺乳動物最早見於晚三疊世,屬始獸類,所見到的化石都是牙齒和頜骨的碎片。
三疊紀時,晚二疊世倖存的齒菊石類大量繁盛起來,中、晚三疊世的大部分菊石有發達的紋飾,
有許多科是三疊紀所特有的。菊石的迅速演化為劃分和對比地層創造了極重要的條件。
雙殼類也有明顯變化,晚古生代的種類只有很少數繼續存在,產生了許多新種類,並且數量相當繁多。
尤其在晚三疊世,一些種屬的結構類型變得複雜,個體也往往比較大。
由於三疊紀的環境與古生代不同,非海相雙殼類逐漸繁盛起來。
裸子植物的蘇鐵、本內蘇鐵、尼爾桑、銀杏及松柏類自三疊紀起迅速發展起來。
其中除本內蘇鐵目始於三疊紀外,其他各類植物均在晚古生代就開始有了發展,但並不占重要地位。
二疊紀的乾燥性氣候延續到了早、中三疊世,到了中三疊世晚期植物才開始逐漸繁盛。
晚三疊世時,裸子植物真正成了大陸植物的主要統治者。
 |
 |
 |
 |
 |
 |
| 雙齒旱魚(Dicellopyge) | 鳥鱷(Riljasuchus) | 松柏木材(Conifer Wood) | 箭齒龍(Belodon) | 茅臺混魚龍(Mixosaurus) | 椰子魚(Birgeria) |
侏羅紀 爬行動物和裸子植物的時代
侏羅紀(Jurassic period)是中生代的第二個紀,始於距今2.03億年,結束於1.35億年,共經歷了6800萬年。
生物發展史上出現了一些重要事件,引人注意。
如恐龍成為陸地的統治者,翼龍類和鳥類出現,哺乳動物開始發展等等。
陸生的裸子植物發展到極盛期。淡水無脊椎動物的雙殼類、腹足類、葉肢介、介形蟲及昆蟲迅速發展。
海生的菊石、雙殼類、箭石仍為重要成員,六射珊瑚從三疊紀到侏羅紀的變化很小。
棘皮動物的海膽自侏羅紀開始佔領了重要地位。
侏羅紀時爬行動物迅速發展。槽齒類絕滅,海生的幻龍類也絕滅了。
恐龍的進化類型——鳥臀類的四個主要類型中有兩個繁盛於侏羅紀,飛行的爬行動物第一次滑翔於天空之中。
鳥類首次出現,這是動物生命史上的重要變革之一。
恐龍的另一類型——蜥臀類在侏羅紀有兩類最為繁盛:一類是食肉的恐龍,另一類是笨重的植食恐龍。
海生的爬行類中主要是魚龍及蛇頸龍,它們成為海洋環境中不可忽視的成員。
三疊紀晚期出現的一部分最原始的哺乳動物在侏羅紀晚期已瀕於絕滅。
早侏羅世新產生了哺乳動物的另一些早期類型——多瘤齒獸類,它被認為是植食的類型,至新生代早期絕滅。
而中侏羅世出現的古獸類一般被認為是有袋類和有胎盤哺乳動物的祖先。
軟骨硬鱗魚類在侏羅紀已開始衰退,被全骨魚代替。
發現於三疊紀的最早的真骨魚類,到了侏羅紀晚期才有了較大發展,數量增多,但種類較少。
侏羅紀的菊石更為進化,主要表現在縫合線的複雜化上,殼飾和殼形也日趨多樣化,
可能是菊石為適應不同海洋環境及多種生活方式所致。
侏羅紀的海相雙殼類很豐富,非海相雙殼類也迅速發展起來,它們在陸相地層的劃分與對比上起了重要作用。
侏羅紀是裸子植物的極盛期。
蘇鐵類和銀杏類的發展達到了高峰,松柏類也占到很重要的地位。
 |
 |
 |
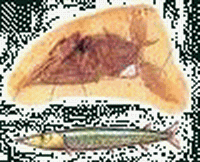 |
 |
 |
| 雲杉(Picea) | 細鱷龍(Compsognathus) | 條棘小嘴貝(Goniorhynchia) | 劍鼻魚(Aspidorhynchus) | 蛇頸龍(Cryptoclidus) | 棱角鱗鱷(Goniopholis) |
白堊紀 爬行動物和裸子植物由極盛走向衰滅
白堊紀(Cretaceus period)是中生代的最後一個紀,始於距今1.35億年,結束於距今6500萬年,其間經歷了7000萬年。
無論是無機界還是有機界在白堊紀都經歷了重要變革。
劇烈的地殼運動和海陸變遷,導致了白堊紀生物界的巨大變化,
中生代許多盛行和佔優勢的門類(如裸子植物、爬行動物、菊石和箭石等)後期相繼衰落和絕滅,
新興的被子植物、鳥類、哺乳動物及腹足類、雙殼類等都有所發展,預示著新的生物演化階段——新生代的來臨。
爬行類從晚侏羅世至早白堊世達到極盛,繼續佔領著海、陸、空。
鳥類繼續進化,其特徵不斷接近現代鳥類。哺乳類略有發展,出現了有袋類和原始有胎盤的真獸類。
魚類已完全的以真骨魚類為主。
白堊紀的海生無脊椎動物最重要的門類仍為菊石綱,菊石在殼體大小、殼形、殼飾和縫合線類型上遠較侏羅紀多樣。
海生的雙殼類、六射珊瑚、有孔蟲等也比較繁盛。
淡水無脊椎動物以軟體動物的雙殼類、腹足類和節肢動物的介形類、葉肢介類為主。
早白堊世仍以裸子植物中的蘇鐵類、本內蘇鐵類、銀杏類和松柏類為主,真蕨類仍然繁盛。
從早白堊世晚期興起的被子植物到晚白堊世得到迅速發展,逐漸取代了裸子植物而居統治地位。
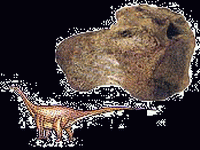 |
 |
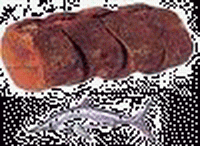 |
 |
 |
 |
| 泰坦龍(Titanosaurus) | 粗板恐龍(Daspletosaurus) | 寬鰭魚(Platyterygius) | 大蓋魚(Macropoma) | 水杉(Malasequoia) | 土豚(Edaphodon) |
被子植物和哺乳類動物時代
第三紀 被子植物的時代
第三紀(Tertiary period)始於距今6500萬年,延至距今180萬年。
第三紀的重要生物類別是被子植物、哺乳動物、鳥類、真骨魚類、雙殼類、腹足類、有孔蟲等,
這與中生代的生物界面貌迥異,標誌著“現代生物時代”的來臨。
第三紀時被子植物極度繁盛。
除松柏類尚占重要地位外其餘的裸子植物均趨衰退。蕨類植物也大大減少且分佈多限於溫暖地區。
第三紀的植物有明顯的分區現象,地層中還有許多微體水生藻類化石。
脊椎動物的變化主要表現在爬行動物的衰亡,哺乳類、鳥類和真骨魚類取而代之,興起且高度繁盛。
第三紀的早期,仍生活著古老、原始的哺乳動物;
到了中期,現代哺乳動物的祖先先後出現,逐漸代替了古老、原始的哺乳動物;
第三紀晚期,現代哺乳動物群逐漸形成,更是偶蹄類和長鼻類繁盛的時期。
尤其馬的進化很快。
中生代末,海生無脊椎動物有明顯的興衰現象。
盛極一時的菊石類完全絕滅,箭石類極度衰退,而雙殼類、腹足類、有孔蟲、六射珊瑚、海膽、苔蘚蟲等則進一步繁盛。
第三紀出現的有孔蟲分佈廣泛、進化迅速,對於海相第三系的劃分與對比很有意義。
此外,它們的生態分帶已應用於確定沉積盆地水深的變化。
原生動物中的放射蟲在第三紀也十分繁盛,在深海研究中佔有突出地位。
雙殼類在第三紀有很大發展,腹足類在第三紀進入極盛期。
陸生的無脊椎動物以雙殼類、腹足類、介形類為主,可以根據它們不同時期組合面貌的變化,進行陸相第三系的劃分
 |
 |
 |
 |
 |
 |
| 櫟屬(Cuercus) | 恐象(Deinolherium) | 乳齒象(Mammut) | 棕櫚木(Palmoxylon) | 榕屬(Ficus) | 背脊鯨(Basilosaurus) |
第四紀 勞動創造了人類
第四紀(Quaternary period)是地球歷史的最新階段,始於距今175萬年。
第四紀包括更新世和全新世兩個階段,二者的分界以地球上最近一次冰期結束、氣候轉暖為標誌,大約在距今1萬年前後。
第四紀生物界的面貌已很接近於現代。
哺乳動物的進化在此階段最為明顯,而人類的出現與進化則更是第四紀最重要的事件之一。
哺乳動物在第四紀期間的進化主要表現在屬種而不是大的類別更新上。
第四紀前一階段——更新世早期哺乳類仍以偶蹄類、長鼻類與新食肉類等的繁盛、發展為特徵,
與第三紀的區別在於出現了真象、真馬、真牛。更新世晚期哺乳動物的一些類別和不少屬種相繼衰亡或滅絕。
到了第四紀的後一階段——全新世,哺乳動物的面貌已和現代基本一致。
大量的化石資料證明人類是由古猿進化而來的。
古猿與最早的人之間的根本區別在於人能製造工具,特別是製造石器。
從製造工具開始的勞動使人類根本區別於其他一切動物,勞動創造了人類。
另一個主要特點是人能直立行走。從古猿開始向人的方向發展的時間,一般認為至少在1000?萬年以前。
第四紀的海生無脊椎動物仍以雙殼類、腹足類、小型有孔蟲、六射珊瑚等占主要地位。
陸生無脊椎動物仍以雙殼類、腹足類、介形類為主。其他脊椎動物中真骨魚類和鳥類繼續繁盛,兩棲類和爬行類變化不大。
高等陸生植物的面貌在第四紀中期以後已與現代基本一致。
由於冰期和間冰期的交替變化,逐漸形成今天的寒帶、溫帶、亞熱帶和熱帶植物群。
微體和超微的浮游鈣藻對海相地層的劃分與對比仍十分重要。
 |
 |
 |
 |
 |
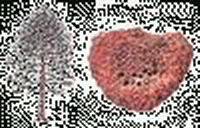 |
| 智慧人(Homo Sapiens Sapiens) | 泥安德特人(Homo Sapiens Neanderthalensis) | 居住人(Homo Habilis) | 巨型猿人(Gigantopithecus) | 澳洲猿人(Australopithecus) | 南洋杉(Araucaria) |
http://museum.pku.edu.cn/exp/paleo/web_his/web/web_hisbe.asp 北京大學地質數字博物館